Family Caesalpinioideae (Leguminosae-Caesalpinioideae)
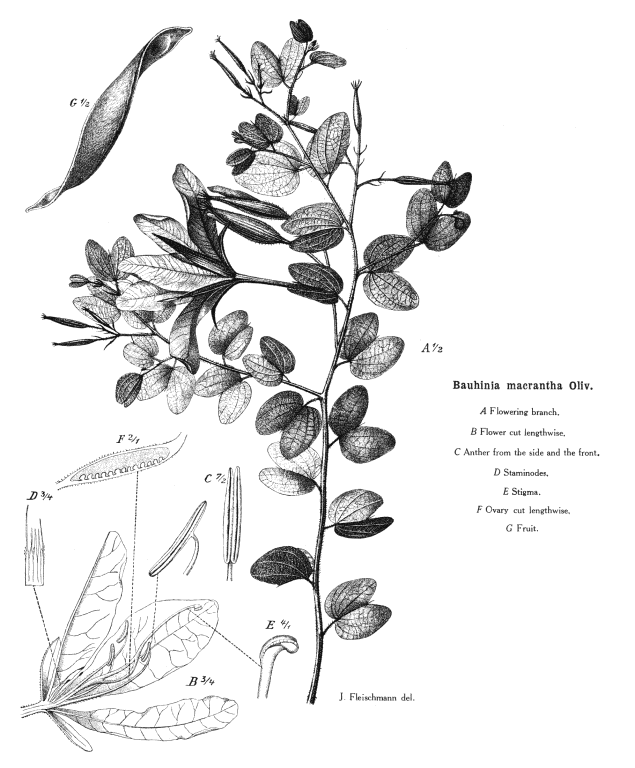
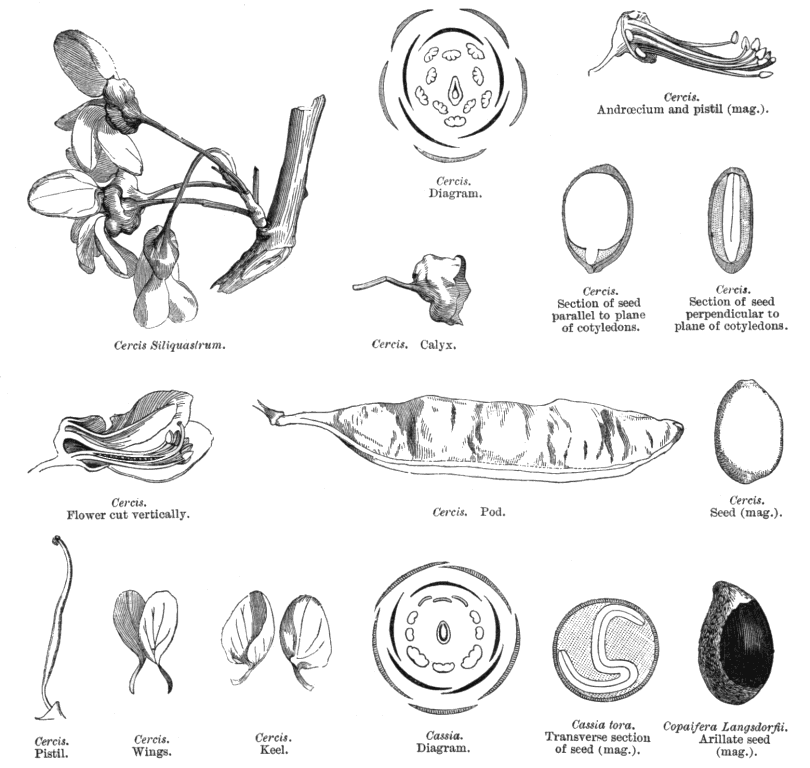
Leguminosae-Caesalpinioideae KunthAlternatively Caesalpiniaceae R.Br.~ Leguminosae Including Ceratoniaceae Link, Detariaceae (DC.) Hess Habit and leaf form. Trees, or shrubs, or herbs, or lianas; resinous, or not resinous. ‘Normal’ plants (mostly), or switch-plants; the switch forms with the principal photosynthesizing function transferred to stems, or phyllodineous. Leaves well developed (usually), or much reduced. The herbs perennial; with neither basal nor terminal aggregations of leaves. Self supporting (mostly), or climbing; the climbers stem twiners, or tendril climbers (via stem or leaf tendrils), or scrambling. Mesophytic, or xerophytic. Heterophyllous (rarely), or not heterophyllous. Leaves evergreen, or deciduous; minute to very large; alternate; spiral, or distichous; ‘herbaceous’, or leathery, or membranous, or modified into spines; petiolate to subsessile (usually), or sessile; non-sheathing; gland-dotted, or not gland-dotted; compound (commonly), or simple (or ostensibly so); pulvinate (usually), or epulvinate; when compound, unifoliolate, or ternate, or pinnate (commonly, either pari- or imparipinnate), or bipinnate (commonly), or bifoliolate (e.g. some Bauhinieae). Leaflets pulvinate, or epulvinate. Lamina when simple dissected (bilobed in some Bauhinieae), or entire. Leaves stipulate (nearly always?), or exstipulate (?). Stipules intrapetiolar; scaly, or leafy, or spiny; caducous, or persistent. Leaves without a persistent basal meristem. Leaf anatomy. Mucilaginous epidermis present, or absent (?). Stomata paracytic, or anomocytic, cyclocytic, and actinocytic. Lamina dorsiventral; with secretory cavities, or without secretory cavities. Secretory cavities containing oil, or containing mucilage, or containing resin (?). The mesophyll containing mucilage cells, or not containing mucilage cells (?); with sclerencymatous idioblasts, or without sclerenchymatous idioblasts. Minor leaf veins with phloem transfer cells, or without phloem transfer cells (for taxonomic details, see Watson and Gunning 1981). Stem anatomy. Secretory cavities present, or absent. Cork cambium present; initially deep-seated, or superficial (?). Nodes tri-lacunar, or penta-lacunar (?). Primary vascular tissue in a cylinder, without separate bundles, or comprising a ring of bundles (?). Medullary bundles absent. Internal phloem absent. Secondary thickening developing from a conventional cambial ring, or anomalous; when anomalous, via concentric cambia (e.g. Koompassia). The secondary phloem stratified into hard (fibrous) and soft (parenchymatous) zones, or not stratified (?). ‘Included’ phloem present, or absent (?). Xylem with vessels. Vessel end-walls simple. Vessels with vestured pits, or without vestured pits (?). Wood storied, or partially storied (VPI); parenchyma apotracheal, or paratracheal (?). Sieve-tube plastids P-type; type IV (subtype (b) in 18 genera). Reproductive type, pollination. Plants hermaphrodite, or monoecious, or andromonoecious, or polygamomonoecious. Pollination mechanism conspicuously specialized, or unspecialized. Inflorescence, floral, fruit and seed morphology. Flowers solitary, or aggregated in ‘inflorescences’; in racemes, or in corymbs, or in fascicles, or in panicles. The ultimate inflorescence unit racemose. Inflorescences terminal, or axillary. Flowers minute to large; somewhat irregular to very irregular (mostly), or regular (not infrequently); commonly more or less zygomorphic; resupinate (?), or not resupinate. The floral irregularity involving the perianth and involving the androecium. Flowers ‘pseudo-papilionaceous’ (usually, the corolla ‘ascending’), or papilionaceous (corolla ‘descending’ in some Cassieae), or neither papilionaceous nor pseudo-papilionaceous (the corolla being at least sometimes absent in 26 genera); pentamerous or not; tricyclic, or tetracyclic (usually), or pentacyclic to polycyclic. Floral receptacle developing a gynophore (often), or with neither androphore nor gynophore. Free hypanthium present (e.g. in most Detarieae), or absent. Perianth with distinct calyx and corolla (mostly), or sepaline; (3–)5, or (6–)10(–11); (1–)2 whorled; isomerous, or anisomerous. Calyx 5, or (3–)5(–6); 1 whorled; polysepalous, or partially gamosepalous, or gamosepalous (mostly); unequal but not bilabiate, or bilabiate, or regular; imbricate, or valvate; with the median member anterior. Corolla (1–)5; 1 whorled; polypetalous, or gamopetalous (in Sympetalandra only); imbricate (always, and usually ascending); green, or white, or yellow, or orange, or red, or pink, or purple. Petals clawed, or sessile. Androecium (1–)10, or 10–50 (often ten, but commonly fewer, ‘many’ in Maniltoa). Androecial members unbranched; free of the perianth; all equal, or markedly unequal; free of one another (sometimes), or coherent (in a variety of configurations); when cohering 1 adelphous, or 2 adelphous; 1 whorled. Androecium exclusively of fertile stamens, or including staminodes. Stamens (1–)3–10(–50); reduced in number relative to the adjacent perianth, or isomerous with the perianth, or diplostemonous to polystemonous. Anthers separate from one another, or connivent; dorsifixed, or basifixed; versatile, or non-versatile; dehiscing via pores (some Cassieae), or dehiscing via longitudinal slits; latrorse to introrse; tetrasporangiate. Endothecium developing fibrous thickenings. Microsporogenesis simultaneous. Anther wall initially with one middle layer, or initially with more than one middle layer (?); of the ‘dicot’ type. Tapetum usually glandular. Pollen shed in aggregates (very rarely — Afzelia, Diptychandra), or shed as single grains; when aggregated, in tetrads. Pollen grains aperturate; (2–)3(–4) aperturate, or 6 aperturate (?); colporate (commonly), or porate, or colpate, or rugate (?); 2-celled (11 genera). Gynoecium 1 carpelled. Carpels reduced in number relative to the perianth. The pistil 1 celled. Gynoecium monomerous; of one carpel; superior. Carpel apically stigmatic; 1–100 ovuled (i.e. to ‘many’, usually in alternating rows along the placenta). Placentation marginal (along the ventral suture). Gynoecium median (the placenta posterior, on the ventral suture). Ovary sessile to stipitate. Ovules pendulous to ascending (?); biseriate; arillate, or non-arillate; anatropous, or campylotropous to amphitropous, or hemianatropous (?); bitegmic; crassinucellate. Outer integument contributing to the micropyle, or not contributing to the micropyle. Embryo-sac development Polygonum-type. Polar nuclei fusing prior to fertilization. Antipodal cells formed; 3; not proliferating; ephemeral, or persistent (mostly). Synergids hooked. Endosperm formation nuclear. Endosperm haustoria present; chalazal, or lateral (rarely). Embryogeny onagrad, or asterad, or caryophyllad (?). Fruit non-fleshy, or fleshy. The fruiting carpel dehiscent, or indehiscent; a legume (usually), or a follicle, or an achene, or samaroid, or a loment, or drupaceous. Fruit elastically dehiscent, or passively dehiscent (?). Dispersal unit the seed, or the fruit. Seeds endospermic (sometimes copiously so), or non-endospermic; small to large (?). Seeds with starch, or without starch (?). Seeds with amyloid, or without amyloid. Cotyledons 2; flat. Embryo chlorophyllous; straight (mostly), or curved (radicle usually straight, rarely somewhat oblique). Micropyle zigzag, or not zigzag (?). Seedling. Germination phanerocotylar, or cryptocotylar. Physiology, biochemistry. Nitrogen-fixing root nodules present (seemingly rarely, by contrast with the other subfamilies), or absent. Cyanogenic, or not cyanogenic (?). Cynogenic constituents tyrosine-derived, or phenylalanine-derived, or of Hegnauer’s ‘Group C’, or leucine-derived (?). Alkaloids present (commonly), or absent (?). Iridoids not detected. Proanthocyanidins present, or absent (?); when present, cyanidin, or delphinidin, or cyanidin and delphinidin (?). Flavonols present (mostly), or absent (?); kaempferol and quercetin, or quercetin and myricetin, or kaempferol, quercetin, and myricetin (?). Ellagic acid consistently absent. Arbutin present, or absent. Aluminium accumulation not found. Sugars transported as sucrose (in all 15 genera sampled). C3. C3 physiology recorded directly in Acacia, Cassia, Cercidium, Gleditsea, Hoffmanseggia. Anatomy non-C4 type (Acacia, Cassia, Mimosa, etc. — all and 150 genera examined by L.W., see Watson and Dallwitz 1983). Geography, cytology. Paleotropical, Neotropical, Cape, and Australian. Temperate, sub-tropical, and tropical. Mostly tropical and subtropical. Taxonomy. Subclass Dicotyledonae; Crassinucelli. Dahlgren’s Superorder Fabiflorae; Fabales. Cronquist’s Subclass Rosidae; Fabales. APG 3 core angiosperms; core eudicot; Superorder Rosanae; fabid; Order Fabales. Species about 400 (?). Genera about 166; Acrocarpus, Adenolobus, Afzelia, Amherstia, Androcalymma, Anthonotha, Apaloxylon, Aphanocalyx, Aprevalia, Apuleia, Arapatiella, Arcoa, Augouardia, Baikiaea, Barklya, Batesia, Bathiaea, Baudouinia, Bauhinia, Berlinia, Brachycylix, Brachystegia, Bracteolanthus, Brandzeia, Brenierea, Brodriguesia, Brownea, Browneopsis, Burkea, Bussea, Caesalpinia, Campsiandra, Candolleodendron, Cassia, Cenostigma, Ceratonia, Cercidium, Cercis, Chidlowia, Colophospermum, Colvillea, Conzattia, Copaifera, Cordeauxia, Crudia, Cryptosepalum, Cynometra, Daniella, Dansera, Delonix, Detarium, Dialium, Dicorynia, Dicymbe, Didelotia, Dimorphandra, Diptychandra, Distemonanthus, Duparquetia, Eligmocarpus, Elizabetha, Endertia, Englerodendron, Eperua, Erythrophleum, Eurypetalum, Gigasiphon, Gilbertiodendron, Gilletiodendron, Gleditsia, Goniorrachis, Gossweilerodendron, Griffonia, Guibourtia, Gymnocladus, Haematoxylum, Hardwickia, Heterostemon, Hoffmannseggia, Holocalyx, Humboldtia, Hylodendron, Hymenaea, Hymenostegia, Intsia, Isoberlinia, Jacqueshuberia, Julbernardia, Kalappia, Kaoue, Kingiodendron, Koompassia, Labichea, Lasiobema, Lebruniodendron, Lemuropisum, Leonardoxa, Leucostegane, Librevillea, Loesenera, Lophocarpinia, Lysidice, Lysiphyllum, Macrolobium, Maniltoa, Martiodendron, Melanoxylon, Mendoravia, Mezoneuron, Michelsonia, Microberlinia, Moldenhauera, Monopetalanthus, Mora, Neochevalierodendron, Oddoniodendron, Orphanodendron, Oxystigma, Pachyelasma, Paloue, Paloveopsis, Paramacrolobium, Parkinsonia, Pellegriniodendron, Peltogyne, Peltophorum, Petalostylis, Phanera, Phyllocarpus, Piliostigma, Plagiosiphon, Poeppigia, Polystemonanthus, Prioria, Pseudomacrolobium, Pterogyne, Pterolobium, Recordoxylon, Saraca, Schizolobium, Schizoscyphus, Schotia, Sclerolobium, Scorodophloeus, Sindora, Sindoropsis, Stachyothyrsus, Stahlia, Stemonocoleus, Stenodrepanum, Storckiella, Stuhlmannia, Sympetalandra, Tachigalia, Talbotiella, Tamarindus, Tessmannia, Tetraberlinia, Tetrapterocarpon, Thylacanthus, Trachylobium, Uittienia (not Vittienia, see P. van Rijckevorsel in Taxacom, March 2009), Umtiza, Vouacapoua, Wagatea, Zenia, Zenkerella, Zuccagnia. This temporary description reflects incomplete breakdown of esoteric characters across the subfamilies of Leguminosae sensu lato (q.v.). However, it is clear that the many features which tend to distinguish the subfamilies all involve rather numerous exceptions, are very incompletely documented, or are not universally applicable. Illustrations. |
Support our developers





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}